Race, Culture and Human Evolution
From: University of Michigan | By: Rachel CaspariMilford Wolpoff 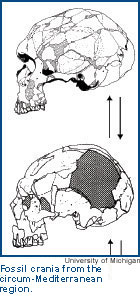 Research on human fossils is exciting; it occurs in exotic places and it stimulates the imagination. But, for the most part, people ponder paleoanthropological questions because the issue is origins, a topic that captures virtually everyone's interest and attention. Paleoanthropology is and has been a public discipline. More than most other science, the study of human evolution deals with issues that people think about and care about. It's our story — our beginnings and our history.
Research on human fossils is exciting; it occurs in exotic places and it stimulates the imagination. But, for the most part, people ponder paleoanthropological questions because the issue is origins, a topic that captures virtually everyone's interest and attention. Paleoanthropology is and has been a public discipline. More than most other science, the study of human evolution deals with issues that people think about and care about. It's our story — our beginnings and our history.
Human evolution also attracts public attention for a more ominous reason. Intrinsic to work on human origins is the recognition of human differences. Part and parcel with human evolution is the study of biological histories and relationships of human populations, the study of what is often considered "race." To understand the genesis of humanity is to grasp the foundation, history, and dynamics of "the races" of humankind.
Race and human evolution are complexly linked, although with common intellectual foundations so deep that the links are often unrecognized. However, in the modern human origins debate of the last decade, the implications of these complex interrelationships for both science and social policy are clear. These implications inspired us to write Race and Human Evolution (1997, Simon and Schuster), a book that examines the historical and modern relationships between race and the study of human prehistory.
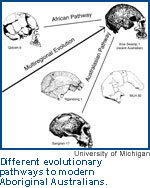
The importance of the relationship between race and human evolution became (sometimes painfully) obvious to us when the multiregional theory that we developed and continue to support entered the domain of public science as the opposition to the popular "Eve" theory. The "Eve" theory suggests that modern humans arose recently, no more than 200,000 years ago, as a new species, and that its "races" then diverged in a bush-like manner as populations spread, separated, and adapted to new regions.
Multiregionalists, in contrast, consider the human species to be much older, one that includes some "archaic" people from many regions of the world. For multiregionalists, human populations have existed in a network of genetic connections, and the "races" they form are temporary channels that diverge and merge in a common evolutionary stream. First introduced by one of us (Wolpoff), with Wu Xinzhi and Alan Thorne in 1984, multiregional evolution was further developed by a number of paleoanthropologists including the other one of us (Caspari), David Frayer, Alan Mann, Geoff Pope, and Dennis Eder, and population geneticists such as Alan Templeton, Simon Easteal, and John Relethford. It is a theory about the ongoing evolutionary process in our species that accounts for the unique pattern of modern human diversity and variation.
The politics of the human origins debate
The politics of the modern human origins debate and its links to race have never been so clear. In the July 27, 1997 New York Times, Chris Stringer and Robin McKie wrote an editorial about newly analyzed Neandertal DNA. (A German team drew ancient DNA from the arm bone of one of the first Neandertals discovered; similar research is ongoing in Andrew Merriwether's laboratory in University of Michigan's anthropology department.) Stringer and McKie took the findings to mean that Neandertal was not an ancestor of living people, and considered this support for the "Eve" theory. They wrote:
The people who replaced the Neanderthals 40 millennia ago, the Cro-Magnons, began to emerge from Africa less than 100,000 years ago. The African emigrants eventually replaced all other hominid species — Neanderthals in Europe, Solo Man in Java and the descendants of Peking Man in China.The implications for the idea of race are profound. If modern humanity is made up of people who are all recent descendants of a few African pioneers, it is equally clear that Homo sapiens must be a startlingly homogenous species. We simply have not had time to diverge genetically in any meaningful manner.
In the past, the races were assumed to be the vestiges of million-year-old cleavages in the human family tree. Race had a profound biological meaning by that reckoning. But now it has become apparent that our differentiation into Eskimos, Bushmen, Australians, Scandinavians and so on occurred only in the last 50,000 years, and that race is a short and superficial coda to the long song of evolution.
Nevertheless, some scientists and those with narrow political agendas have put forward arguments to sustain the idea that races exist with fundamental biological differences.
Over the last decade multiregional evolution has received much popular and scientific attention because of the role it plays as the antithesis to the "Eve" theory of modern human origins, but it is easy to see that this is not always the kind of attention sought. So much heat and intensity about some genes from a dead caveman's arm surely means the issue has poked a sensitive nerve. The nerve does not lie in the dry bones of human evolution, but in interpretations of the most sensitive biological and political topic of this century — human races.
Two explanations for human diversity
Race and human evolution are linked because the two evolutionary theories about humanity's origins are also about its variation. Two elements of human diversity make the pattern of human variation unique among primates. First, unlike any other primate species, humans are widespread, occupying a vast range of ecological niches around the world. Other animal species also occupy wide ranges, of course, but this is almost always because the habitat and ecology they adapt to are broadly distributed. Secondly, in spite of their large range, human populations are each both internally variable and genetically similar; it is said that differences between the most disparate human populations amount to less than what one finds between adjacent populations of a single frog species! More than 85 percent of all uniquely human genetic variation can be found within every human population.
One way to explain low genetic diversity between populations of a species is time: for example, if the species had a very recent origin. According to this explanation, human populations are similar because there hasn't been enough time for differences to develop. This theory requires rapid divergences between populations and is supported by studies that assume a branching pattern of population divergences within Homo sapiens, the same kind of pattern that reflects relationships between species, where there can be no mating between the groups. Genetic differences (or distances) are correlated to time by constructing "molecular clocks" that date the various branch points.
The other explanation of low genetic diversity between populations is constant mixing, or genic exchanges: gene flow between populations and population migrations, creating a network of genetic relationships that can disperse genes to many populations and even throughout the entire human species. Even without the process of natural selection, a low level of genic exchange (one or two individuals each generation) is enough to keep the most far-flung human populations within the same species and to spread neutral genes (genes that neither help nor hinder adaptation). But selection for advantageous features and behaviors diffuses their genes much more rapidly. Multiregionalists recognize that the network-like evolutionary pattern within Homo sapiens cannot be described as a tree because tree branches cannot depict reticulation or merge back together as human populations continually do. The human pattern of genetic diversity reflects the importance of genic exchanges. Human "races" are genetically close because they never were isolated. These races are not branches on an evolutionary tree, and for the most part are not like the races, or subspecies, found in other animals.
Human races
Well, then, what are human races like? Ethnologists such as John Moore of the University of Florida show how to address the question from the perspective that biological, linguistic, and cultural relationships between populations today are multiple and complex. Each of these descends from, or is rooted in, several different antecedent groups. A Parisian might represent a multiplicity of different linguistic and biological backgrounds; she may speak French as a first language but also know Arabic, and she may have biological roots in both North Africa and France. In modern times this is an obvious and ubiquitous observation, but this pattern is not limited to the recent past or to putative "melting pots" like the United States or post-colonial European powers; it is a worldwide phenomenon that also has great time depth. Studies of tribal peoples in the ethnographic present, from the American Great Basin to Australia to Tierra del Fuego, reveal no ethnically isolated groups. Instead, marriages across what appear as the most profound ethnic boundaries are common, and multilingualism prevails.
After working with Pacific Islanders who were once considered isolated, John Terrell of the Field Museum of Natural History now refers to the "Myth of the Primitive Isolate." He argues that biological, cultural, and linguistic similarities between these peoples are due primarily to continuous interactions over time, and are not the simple consequence of common ancestry, as anthropologists have often assumed. Moore calls this "ethnogenesis." Populations, languages, and cultures continue to change, dividing and merging like the channels in a river that can separate and recombine numerous times. A population (or language or culture) can have several ancestors and several descendants. The ethnogenetic perspective is multiregionalism. It interprets human history in terms of interactions across profound ethnic and cultural boundaries, with amalgamation of linguistic traits and recurrent "hybridization" of cultures.
One driving force is the virtual universality of exogamy. It is fundamental to both the networks of genetic connections we described and in the establishment of alliance networks between human groups. These alliances and their roles in complex patterns of competition and cooperation may be the most important element of our social evolution, and one that an increasing number of scientists believe may have driven our biological evolution as well. We expect that socially prescribed patterns of intergroup genic exchange were important for as long as humans were cultural and communicated with a complex language. Evolutionary models that depict human groups as isolated branches on a tree are more than overly simplistic — they portray an evolutionary pattern for our species that does not exist and has not existed.
The implications for human races are profound. In mammalian systematics, "race" and "subspecies" usually mean much the same thing. These terms describe genetically different, geographically defined groups of populations that contrast enough with each other for a relatively small number of characters to distinguish the individuals of one from another. Chimpanzees, even in their limited range across equatorial Africa, are divided into at least four subspecies, with far more variation between them, and less common variation within them, than are human races, no matter how they are defined.
Human races are nothing like this. They are far less different from one another, and they are not distinct lines of descent, because each has multiple ancestors. This is not to say that the human species is homogeneous, or that its features are uniformly distributed. There is geographic variation within our species, and within broad regions it is possible to find some common combinations of physical features that reflect physical proximity, common adaptation, and ancestral geographic identity. But this is not much like they pattern of how features are distributed within the subspecies of most other species. While "race" can more clearly describe geographic variation in many other species, there are significant problems in using the term to describe human geographic variation, because of the pattern of mixing we describe above and since the primary meaning of "race" is sociocultural and is only very loosely linked to biology.
Hence, the American racial category "Black" may include people whose ancestry is from Africa, parts of India, or even the New Guinea Highlands! Only a few years ago, in apartheid South Africa, Japanese were considered "white," while Chinese were considered "colored." These "races" are clearly cultural constructions, yet as racial classifications they are treated by society as though they were grounded in biology. The problem is twofold: the indistinct nature of human variation we described, and the agendas that come into play whenever categories are cultural constructs. Some years ago we heard an apocryphal story of an American reporter interviewing "Papa Doc" Duvalier, then dictator of Haiti. Race came up in the conversation and the reporter asked Papa Doc: "How many Haitians are white?"
"Oh, close to 95 percent," he replied.
The visibly surprised reporter questioned this, asking, "But how do you define white?" Papa Doc answered, "How do you Americans define black?"
When the reporter smugly responded, "That's easy, anybody with some African ancestry," Papa Doc agreed, "We define white using the same principle."
Geographic variation exists, of course, and we often incorporate it into our social-racial constructs. But this variation is a dynamic and ever-changing consequence of human demography and evolution that reflects many factors except separate lines of descent and is broadly explained by multiregional evolution. Unfortunately, anthropologists have not agreed on a more appropriate word than "racial" for this form of variation, and the use of the term remains controversial.
Multiregional evolution
 Multiregional evolution was developed to address the question of geographic variation and its temporal dimension and how geographic variation fits into the picture of a widespread evolving species. Threads of common adaptation differing from one place to another are easy to understand, as long as the environment does not fluctuate too much. Short, stocky bodies are part of a cold adaptation, and can either persist or appear again and again whenever human populations adapt to cold environments. But there is something of a paradox in how historically influenced variation can persist in spite of ongoing genetic exchanges. What accounts for long-lasting regional and populational differences, such as the coalshovel-like inner surface of the incisors of Asian peoples, if not genetic isolation? What accounts for common evolutionary changes across a widely scattered species, such as the increases in brain size that happen everywhere, if not genic exchanges dispersing new adaptations?
Multiregional evolution was developed to address the question of geographic variation and its temporal dimension and how geographic variation fits into the picture of a widespread evolving species. Threads of common adaptation differing from one place to another are easy to understand, as long as the environment does not fluctuate too much. Short, stocky bodies are part of a cold adaptation, and can either persist or appear again and again whenever human populations adapt to cold environments. But there is something of a paradox in how historically influenced variation can persist in spite of ongoing genetic exchanges. What accounts for long-lasting regional and populational differences, such as the coalshovel-like inner surface of the incisors of Asian peoples, if not genetic isolation? What accounts for common evolutionary changes across a widely scattered species, such as the increases in brain size that happen everywhere, if not genic exchanges dispersing new adaptations?
The answer involves the interactions between the evolutionary forces: selection (genetic difference from having different numbers of surviving offspring), genetic drift (random change because of small samples), and, most importantly, genetic exchanges (dispersal of genes by population movements or because of mate exchanges). A role for exchanging genes to promote differences seems counter-intuitive, but as discussed below, it doesn't just account for similarities between populations. There are two other important elements: isolation by distance (continuous variation from the source of a new gene to its furthest occurrence, which can be great enough to act as though populations across an unbroken range were actually isolated), and clinal balances (continuous variation created when opposing evolutionary forces act at different parts of a species range). These insure that genic exchanges not only spread favorable genes but also contribute to the persistence of differences between groups. The actions of these forces, influenced by human culture, account for the unusual pattern of biological variation found in the human species.
To some extent, the geographic distribution of human variation reflects the immediate consequences of the dispersals and population movements in the Early Pleistocene (especially between one and one-half and three-quarters of a million years ago), as humanity transformed into a colonizing species, and there were significant population movements out of Africa. During the colonization process, as ranges extended into uninhabited territories, populations separated and dispersed widely. The initial colonizing populations were quite small, and we assume that many of the colonizations were unsuccessful. Fluctuating in size and often splitting, each surviving group had a unique history in which different features were lost or became common because of genetic drift.
As population sizes grew and people increasingly came into contact, their genetic exchange network became more complex. Populations continued to merge, and split again, as ethnogenic processes similar to those of today were established. Geographic differences developed, with contrasts between the larger, heterogeneous populations at the species center and place of origin, and smaller, more homogeneous but differently distinct populations toward its edges. The fact that many more people lived at the center for most of human evolution, which for humanity is Africa, promoted this pattern: genic exchanges were greatest here. At the edges, populations had less contact with one another, and there the evolutionary forces of selection and drift were often the strongest. One consequence of this is that in these peripheral regions, some of the features that mark modern geographic variation have been found to appear in the earliest immigrants.
Now we return to the paradox: even the most outlying human populations were not genetically isolated, yet some regional distinctions are maintained for very long time periods. Most once-common features do disappear with time, but fossil evidence suggests that some can be very long lasting. Why? One reason is that when a feature is advantageous, selection helps it persist; some features may be under selective pressure that doesn't change. Other features may become incorporated into new adaptations as selection changes; they become exaptations, adaptive features that did not originate as part of their current adaptation. Another reason for this persistence reflects a kind of evolutionary momentum. Features that no longer improve adaptation because environmental factors have changed, or nonadaptive features initially established at high frequencies due to drift, may remain unaltered as long as no evolutionary forces act to change them.
Social behaviors may provide an important source of long-standing selection. Facial features can underscore or reinforce linguistic and other cultural elements that influence mate choice and also may be indicators of social identity and important in kin recognition. Recognition of kin comes to play a double role in humans, where social systems of kinship and alliance play key parts, not only in mate choice but also in other interactions where shared genes are important. Behaviors that extend aid to related individuals can help one promote one's own genes, because relatives are more likely to have some of the same genes than unrelated individuals are. Many scientists have suggested that human social evolution is influenced by the special behaviors of individuals toward their kin. Human society helps related individuals help each other, for instance by defining groups of related individuals.
But how can we tell who is related to us? There may be cultural clues: clothing, jewelry, or special mannerisms or words. Biology can also be important, and visual recognition of related individuals is largely based on features of the face. Humans have neurological adaptations to help obtain the maximum amount of information for such identifications. It is interesting that most people derive clues about origin from these features. Ideas about "self" and "other" are partially based on physical features in many, if not most, societies around the world. It is almost certainly not a coincidence that the face is where many of the externally visible features showing regional continuity are found, as facial characteristics play an important role in helping to recognize those related by systems of kinship or alliance.
So facial and some other features may have been maintained by selection, but they were often first established by genetic drift. Paradoxically, drift may mimic the role of selection in creating difference between populations, maintaining characteristics common in one region or another, especially toward the peripheries. As the most remote regions across the human range were colonized, populations there were subject to harsher conditions and climatic fluctuations of greater magnitudes than at the center. Numerous local population extinctions and recolonizations from nearby created a population structure in which broad regional continuity could be expected even though many local populations died out, because it was usually pioneers from nearby, genetically similar populations who recolonized the areas of extinct populations. This kind of population structure allowed drift to continue playing a significant role in regional evolution, while there was long-lasting regional continuity for some features.
Clinal variation
The features that demonstrate regional continuity over time are not unique to any particular region; while they are found one place or another in high frequency, most have a continuous geographic distribution. Many of the most obvious and interesting human characteristics such as body shape and skin color vary systematically over space. Often these gradients are consequences of varying selection pressures discussed below. However, this variation can also occur in selectively neutral traits like the surface anatomy of teeth, due to isolation by distance, a concept first understood and modeled by Sewall Wright over a half-century ago. Even if populations had been homogeneous from the onset (which they were not), differences between them would be inevitable, especially as humans came to occupy different habitats. Gene flow alone will cause neutral genes to spread, but populations are distributed multidimensionally; genes take time to disperse over great distances, and have a better chance of getting lost where populations are sparse and population sizes are small. For this reason the neutral genes will not be uniformly distributed and population structure (the opposite of population homogeneity) is created.
A second cause of genetic variation over space was recognized by the mathematician Per Enflo. He noted that human populations differed (as they still differ) in what he calls their reproductive stability. Some maintain constant population sizes for long time periods, while others fluctuate in response to diseases, or ecological or climatic changes. People will tend to move from reproductively stable to unstable regions, which can lead to some genes dispersing widely but not necessarily evenly. Selection also can distribute features across broad gradients of variation when the features are not neutral. For instance, skin color corresponds to the amount of solar radiation that skin is exposed to, and it varies along with the maximum exposure to sunlight. When the source of selection varies gradually over space, as maximum solar radiation does, the adaptive response varies as well and geographic distinctions can directly correspond to selective pressures. This is why skin color can be seen to follow a continuous gradient from north to south in both Old and New World populations.
 However, there is another, more complex source of clinal variation that explains how regional features can be maintained even when advantageous features disperse. This is when there is a gradient formed between opposing causes of evolution — genic exchanges between adjacent populations vs. selection or drift. These can interact to create a pattern of continuous variation. Population movements are often from central, more densely occupied areas to more sparsely inhabited peripheral regions with greater reproductive instabilities. Advantageous new genes or gene combinations may first appear at or near the center, where there are more people, better adaptation, and higher reproductive stability, and then spread outward. The dispersal may reflect differences in population size, reproductive stability, or advantages conferred by the genes. Advantageous genes can also appear in peripheral regions, of course, and disperse to the center and other peripheries because of selection. Toward the peripheries, however, there may be opposing evolutionary forces, perhaps selection against the innovation, or significant loss of new genes because peripheral populations are very small and accidental changes can be important. This will create a gradient of continuous variation.
However, there is another, more complex source of clinal variation that explains how regional features can be maintained even when advantageous features disperse. This is when there is a gradient formed between opposing causes of evolution — genic exchanges between adjacent populations vs. selection or drift. These can interact to create a pattern of continuous variation. Population movements are often from central, more densely occupied areas to more sparsely inhabited peripheral regions with greater reproductive instabilities. Advantageous new genes or gene combinations may first appear at or near the center, where there are more people, better adaptation, and higher reproductive stability, and then spread outward. The dispersal may reflect differences in population size, reproductive stability, or advantages conferred by the genes. Advantageous genes can also appear in peripheral regions, of course, and disperse to the center and other peripheries because of selection. Toward the peripheries, however, there may be opposing evolutionary forces, perhaps selection against the innovation, or significant loss of new genes because peripheral populations are very small and accidental changes can be important. This will create a gradient of continuous variation.
Clinal balances are independent of the absolute magnitude of the forces and depend only on their relative sizes. The multiregional model is based on the proposal that these different kinds of continuous variation and dynamic clinal balances are the causes of human biological variation today, and were important throughout Pleistocene human evolution. Three lines of evidence in the fossil record indicate that this pattern has great antiquity, extending to the earliest human colonizations outside of Africa. The first is evidence that during the colonization period of human evolution, all humans belonged to a single species, possessing great behavioral flexibility permitted by a common cultural capacity. The second is the observation of regional continuity, which undermines arguments for a recent speciation by showing that complete replacements throughout all of the human range could not have happened. Therefore, these mechanisms account for observations of both regional continuity and common evolutionary change, and provide a historical as well as a spatial dimension to modern population variation. The third is the recognition, based on anatomical and archaeological evidence, that no region was ever isolated for any significant time. Common features and tools continue to disperse throughout the human range in every period of human prehistory.
The importance of Culture
There are two very distinct parts to human evolutionary history, and the general patterns are quite different for each. In the longer part, for the first two-thirds of our evolution, humans were geographically restricted and their evolutionary pattern was similar to that of other primate species, most of which are adapted to limited ecological niches. Adaptive variation was not found within one hominid species, but between different, competing ones.
In the second shorter and more recent colonization phase, a single species of humans came to exist all over the Old World. Beginning almost two million years ago, people with longer legs and brains that were substantially larger and much more energy consuming than those of their ancestors were the first to begin colonizing the world outside of Africa. These people possessed behaviors that allowed them to establish themselves in numerous environments, and they are known from fossil sites all over the Old World. In the 1960s, the taxon "Homo erectus" came to be used for these Middle Pleistocene humans, replacing a plethora of generic and specific names used to represent geographic variants of the taxon. Virtually all students of human evolution recognized that in spite of geographic dispersion, there was only one species.
In recent years, though, there has been a tendency for some scientists to consider geographic variants of Middle Pleistocene hominids to be different species, geographic populations that did not and could not interbreed. These determinations are based on minor anatomical differences, sometimes the same evidence for regional continuity we have been discussing, but interpreted in a different theoretical framework than ours. This different framework does not treat geographic groups as evolving, interconnected elements in a single behavioral and genetic network that incorporates variation as a natural attribute, but rather interprets geographic groups as different branches on a tree. Criteria are sought, no matter how minor, to prove the variations reflect different species that can be distinguished. These criteria involve minute differences between the branches and are similar to those that were applied to categorize modern human variation a century ago, used to "demonstrate" that human races were different species.
The second part of our evolutionary history is unusual in a way that addresses this issue. Most geographically dispersed mammals occupy an ecological niche that is broadly distributed, but the human pattern is of a widespread single species with many different ecological niches. The difference is human Culture, the unique human capacity to transform experience into symbols for information storage and transmission and complex communications. Culture allows unprecedented behavioral flexibility, providing critical elements of what is necessary for human populations to occupy so many diverse niches. Culture affects evolutionary processes in many other respects; it influences the demography of populations and plays an inestimably important role in shaping their evolutionary histories. While it is unlikely that culture appeared suddenly in its present form — its definition and evolution are obviously complex — the behavioral flexibility it allows was clearly significant long ago, during the colonizing phase of human evolution. This is why humans in this later phase did not follow the species proliferation and extinction patterns of other mammals that lack this degree of flexibility. Moreover, it is likely that the cultural behaviors that promoted geographic expansion also contributed to multiregional evolutionary mechanisms.
It has been suggested that human "intelligence" and brain expansion occurred as a consequence of social evolution that involved the creation of large-scale alliance networks used in social strategies involving both competition and cooperation between groups. These relationships necessitated complicated neural mapping afforded by a large and restructured brain, because they depend on expectations that are not necessarily shared, but are always mutually interpretable. Alliances are usually created and reinforced through marriage systems. Thus, exogamous social institutions that foster genic exchanges likely played an important part in the evolution of culture and of the human brain.
Humans that first left Africa had brain sizes within the modern range of variation, although considerably lower than the average modern brain size. Their artifacts indicate that they all shared a similar cultural capacity, and they were socially and technologically capable of living in harsh environments. It is probable that they, like modern humans, possessed cultural behaviors that facilitated genetic exchange. Population geneticists such as Alan Templeton have shown that relatively low levels of exogamy could maintain the genetic continuity across geographic regions required by the multiregional model. In fact, Templeton's research on mitochondrial and nuclear DNA shows that these exchanges have been going on for a very long time, because they left their mark on the distribution of genes today.
Human variation is and always has been complex. Branches on an evolutionary tree do not describe the histories and relationships of modern populations, nor do they reflect those of the past. Yet those branches reflect popular conceptions of races as distinct "types" with separate evolutionary histories. With roots in 19th-century science, these conceptions have permeated society as a whole, a society that in turn contributes to human evolutionary theory by influencing scientific traditions. Thus, for Stringer and McKie, those scientists who disagree with their "Eve" theory must be racist. For researchers who assume that a branching pattern defines relationships between human populations, great antiquity of the human species means very long branches indeed, but it is the short branches of their theory that are a symbol of brotherhood. These preconceptions create analyses of population difference that start with the assumptions of divergence and isolation. But trees do not describe the evolutionary process within Homo sapiens, a process that is best explained by multiregional evolution. The multiregional perspective shows how Darwinian evolution can create human differences without isolation and divergence, and can account for the unique pattern of diversity found in Homo sapiens.